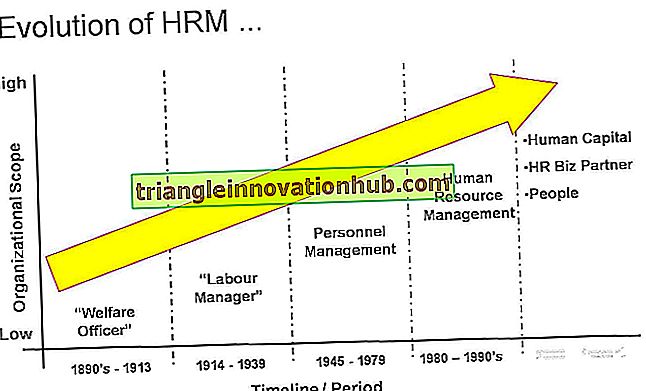
Merits and Demerits of Telome Theory
Enkele van de verdiensten en nadelen van de Telome-theorie zijn als volgt:
De interpretatie van het plantenlichaam als een vertakte as met zijn afdelingen die zijn gespecialiseerd voor verschillende functies, maar allemaal zoals in de fundamentele structuur, wordt de telome-theorie genoemd. De telome-theorie is te danken aan het werk van Zimmermann.

Afbeelding Courtesy: upload.wikimedia.org/wikipedia/commons/7/71/Athyrium_filix-femina.jpg

Hij keert terug naar het vroegste en eenvoudigste type van de bekende vaatplant, namelijk in de Psilophytales waarin het lichaam bestaat uit vertakte, bladloze assen, die allemaal in wezen gelijk zijn. In sommige gevallen kunnen deze dichotomiëel in andere monopodiaal worden gerangschikt, en het verschil kan zijn ontstaan door 'overtopping', maar in beide gevallen zijn deze naakte assen de primitieve bouweenheden van de cormophyten, die dus in de eerste plaats als een assenstelsel worden beschouwd .

Elk van de terminale vertakkingen in een dergelijk systeem wordt een telome genoemd. Elk telome is een ultieme vertakking op een oudere as of mesoom. Sommige telomes zijn steriel; andere dragen terminale sporangia en zijn daarom vruchtbaar. Telomes hebben ook de neiging zich te verenigen in groepen, genaamd, syntelomes, die allemaal steriel, allemaal vruchtbaar of gemengd kunnen zijn.
Volgens de telome-theorie zijn de vroegste dijken afgeplatte telomes of phylloïden, en het sporangium is in alle gevallen, samen met zijn steel of sporangiofoor, een vruchtbaar teloom. Samengestelde sporangioforen, zoals die van Equisetum, zijn syntelomes. De phylloïden zijn de prototypen van de microphyllene bladblaadjes.
In de meest primitieve omstandigheden waren de steriele en vruchtbare telomes blijkbaar vrij onafhankelijk van elkaar. De axillaire positie die de sporangiale telomen in Lycopsida innemen is bereikt door secundaire verschuiving van de telomen en toont drie manieren waarop de verandering kan hebben plaatsgevonden.

Het plantenlichaam is dus een vertakkende as, zijn gespecialiseerde ondergrondse segmenten zijn wortels zijn luchttoppen, indien vruchtbaar, sporangia, indien steriel, fylloïden. "Bladeren" bestaan onder deze interpretatie uit één of meer steriele telomen die in veel gevallen geassocieerd zijn met één of meer vruchtbare telomes.
Bladmorfologie ondersteunt aldus de verdeling van vasculaire planten in vier hoofdgroepen, de Psilopsida zijn bladloos of tonen bladeren slechts gedeeltelijk gedifferentieerd; het blad van de Lycopsida is vergroot en uitgewerkt; die van de Sphenopsida is een minder belangrijk vertakkingsysteem, dat van Pteropsida, het grootste. Het blad is dus van verschillende aard in vaasvormige planten; de evolutionaire ontwikkeling van een fotosynthetisch orgel is op meer dan één manier tot stand gekomen.

verdiensten:
Dit is een eenvoudig concept en verklaart de meeste morfologische problemen met betrekking tot verschillende organen van een plant. Volgens Bierhorst (1971) is deze theorie te simpel en te gemakkelijk toepasbaar, maar helaas heeft het overmatige gebruik ervan de waarde ervan sterk verminderd.
minpunten:
1. Telome is beschouwd als kant en klare eenheid. Deze moeilijkheid werd gerealiseerd door Zimmermann (1949, 52) en vervolgens herkende hij verschillende andere elementaire processen als (a) interconnectie van cellen; (b) rotatie van de celas; (c) differentiatie van apicale cellen, enz., die hebben geleid tot de vorming van een apicale cel met drie snijvlakken. Deze elementaire processen voldoen echter niet aan de plantenmorfologie (Puri, 1956).

2. Veel andere planten van veel grotere complexiteit dan Rhynia-fossielen zijn ontdekt in bedden van dezelfde leeftijd of zelfs eerder, bijvoorbeeld Zosterophyllum, Baragnnathia (Leclercq, 1954) Lyon Hueber (1964), Hueber en Banks (1967) en Lyon (1964) waargenomen laterale sporangia op korte gevasculariseerde stelen in respectievelijk Psilophyton en Asteroxylon in plaats van gebruikelijke terminale sporangia.
3. Volgens de voorstanders van de telome-theorie zijn alle bladeren in planten telomisch van aard. Enantietheorie daarentegen beschouwt microbladige bladeren als enige uitlopers van de stengel (Bower, 1935).

4. De polystelische conditie in de as van planten wordt verondersteld te zijn ontwikkeld als gevolg van parenchymatische syngenese van verschillende monostatische assen. De actinostelic conditie wordt verondersteld het product te zijn van radiale fusie van steles in de polystelische as. Een dergelijke verklaring druist lijnrecht in tegen het algemeen geaccepteerde concept van de stelar-theorie (Stewart, 1964).
5. Andrews (1963) heeft een reeks diagrammen van paleozoïsche zaden gegeven om de oorsprong van de cupule te verklaren. Pettitt (1970) vond dat ze min of meer van dezelfde leeftijd waren.
6. De theorie heeft weinig aandacht gekregen van angiosperm-gecentreerde morfologen. De toepassing ervan op meeldraden (Puri, 1947, 1951, 1955), het venatiepatroon van bladeren (Foster, 1950), de morfologische aard van angiospermbladeren en sporofylen, en carpellen (Eames 1961) zijn van tijd tot tijd bekritiseerd.
Kortom, het zou de moeite waard zijn om de reactie van Andrews en Eames te citeren. Andrews (1961) zegt: "Zimmermanns plan voor de pteropsiden, of tenminste enkele pteropsids, heeft veel ondersteunend bewijs, zijn concept voor de articulaten kan geldig zijn, maar we staan slechts op het punt om de oorsprong van deze groep te begrijpen, zijn concept voor de lycopsiden zijn, voor zover ik weet, puur hypothetisch. "
Eames (1961) zegt: "beschouwing van het primitieve plantenlichaam, bestaande uit basiseenheden, telomes, is ongetwijfeld waardevol voor het begrijpen van primitievere taxa, maar de waarde ervan in de interpretatie van de hogere taxa, waar as en aanhangsels worden gevestigd als morfologische eenheden, is twijfelachtig. "