Orbit: The Blood Vessels System of the Orbit of Human Eyes (8329 Words)
Hier zijn je aantekeningen over het bloedvatenstelsel van de Orbit of Human Eyes!
Oftalmische slagader:
Het is een tak van de interne halsslagader en ontstaat wanneer dat bloedvat door het dak van de holle sinus komt, mediaal naar het voorste clinoïde proces (figuur 9.20).

Afbeelding Courtesy: newhorizonsnaturalhealthcare.com/linked/cardiovascular.jpg
De ader komt de baan binnen via het optische kanaal infero-lateraal naar de oogzenuw, beide liggen in een gemeenschappelijke omhulling van Dura mater. In de baan doorboort het de Dura, slingert zich langs de laterale zijde van de oogzenuw en passeert voorwaarts en mediaal boven de oogzenuw tussen de superieure oogader aan de voorkant en de nasociliaire zenuw erachter.
Het bereikt de mediale wand van de baan tussen de mediale rectus en superieure schuine spieren, en aan het mediale uiteinde van het bovenste ooglid verdeelt de slagader zich in twee terminale takken, de supra-trochlear en dorsale nasale.

branches:
Takken van de oogader vergezellen alle takken van nasociliaire, frontale en traanzenuwen. Takken zijn gerangschikt in de volgende groepen:
(A) Takken naar de oogbol:
1. Centrale slagader van het netvlies:
Het ontstaat uit de oogheelkundige slagader onder de oogzenuw, loopt vooruit in de durale omhulling en prikt de oogzenuw ongeveer 1, 25 cm achter de oogbol inferomediaal. De ader bereikt de optische schijf door het centrale deel van de zenuw en levert de oogzenuw en de binnenste zes of zeven lagen van het netvlies.
De centrale slagader is een typische end-arterie en de obstructie veroorzaakt totale blindheid.
2. Posterior ciliaire slagaders, bestaan uit twee sets, lang en kort, die beide de sclera rond de oogzenuw doorboren.
Lange posterieure ciliaire slagaders, meestal twee in aantal, bereiken de perifere rand van de iris, anastomose met de voorste ciliaire slagaders van de musculaire takken van de vier recti en vormen een belangrijke arteriële cirkel om de iris en het corpus ciliare te voeden.
Korte posteriore ciliaire slagaders, gewoonlijk zeven in getal aan het begin, breken uiteen in capillaire plexus in de choroïde en leveren de choroïde en avasculaire buitenste drie of vier lagen van de retina door diffusie.
(B) Takken naar de orbitale spieren:
De anterieure ciliaire slagaders zijn afgeleid van de musculaire takken.
(C) Takken langs de zijwand van de baan:
De traanslagader passeert langs de bovenrand van de laterale rectus en levert de traanklier, oogleden en bindvlies. Het biedt twee laterale palpebrale arteriën, één voor elk ooglid, die anastomose met de mediale palpebrale slagaders. Bovendien geeft de traanslagader zygomatische en terugkerende meningeale takken; de laatste passeert de superieure orbitale spleet en anastomose met de middelste meningeale slagaders.
(D) Takken langs de mediale muur van een baan:
1. Laterale ethmoldale slagader, om posterieure ethmoïdale sinussen te leveren;
2. Anterieure ethmoldale slagader, voor het leveren van voorste en middelste ethmoïdale sinussen, antero-superieure delen van de zijwand en het neusvlies;
3. mediale palpebrale arteriën, één voor elk ooglid; elke slagader verdeelt zich in twee takken die zijwaarts langs de boven- en onderranden van de tarsalenplaten buigen.
4. Supra-orbitale en supra-trochleaire aderen vergezellen de overeenkomstige zenuwen en voorzien in het voorhoofd en de hoofdhuid.
5. Dorsale neusslagader levert de externe neus en anastomosen aan de terminale tak van de slagader.
Oftalmische aderen:
Twee aderen draineren de baan, superieure en inferieure oftalmische aderen.
De superieure oogader in het middengedeelte van het bovenste ooglid kruist boven de oogzenuw in samenwerking met de oogader en ontvangt zijrivieren die corresponderen met de takken van de begeleidende slagader.
Het passeert de superieure orbitale spleet en eindigt in de holle sinus. De ader heeft geen kleppen en communiceert bij het begin met de gelaatsader via de hoekader.
De onderste oftalmische ader begint in de bodem van de baan en verzamelt bloed uit de inferieure orbitale spieren, traanzak en oogleden. Het loopt weg in de holle sinus, hetzij direct, hetzij nadat hij is verbonden met de superieure oogader. Het communiceert met de pterygoid veneuze plexus door de inferieure orbitale spleet.
Orbitaal vet:
Het vult het interval tussen de oogzenuw en de kegel van vier recti-spieren. Het werkt als een kussen om de oogbal te stabiliseren.
oogappel:
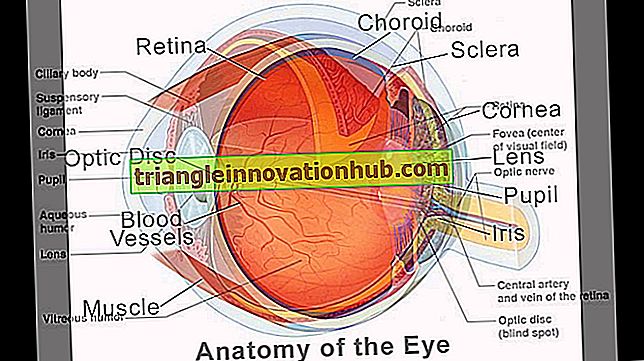
De oogbol of bol van het oog neemt het voorste derde deel van de orbitale holte in en is ingebed in vet gescheiden door een vliezige zak, de fascia bulbi. Het bestaat uit segmenten van twee bollen; de voorste één-zesde die kleinere bol vertegenwoordigt vormt het hoornvlies en achterste vijf-zesde die tot grotere bol behoort, vormt de sclera.
De anterieure en posterieure polen van het oog zijn de centrale punten van de cornea en sclerale krommingen. Een anero-posterieure lijn die beide polen verbindt, vormt de optische as, terwijl een lijn die zich uitstrekt van de voorste pool naar de fovea centralis, die iets lateraal ligt ten opzichte van de achterste pool, de visuele as vormt voor nauwkeurig zicht.
Een denkbeeldige lijn rond de oogbol op gelijke afstand van de twee polen staat bekend als de evenaar. Elk denkbeeldig vlak van pool tot pool en de evenaar onder een rechte hoek snijden, staat bekend als de meridiaan. Daarom kan een meridionaal gedeelte door het oog horizontaal, sagittaal of schuin zijn. De oogzenuw is ongeveer 3 mm aan de neus van de achterste pool bevestigd aan de oogbol.
Elk van de antero-posterior, transversale en verticale diameters van de normale volwassen oogbol meet ongeveer 24 mm. Bij bijziendheid kan de antero-posterieure diameter worden verhoogd tot 29 mm en bij hypermetropie kan deze worden verlaagd tot 20 mm.
De wand van de oogbol, die de brekende media omsluit, bestaat uit drie tunieken of jassen. De buitenste tuniek is vezelig en bestaat uit sclera en hoornvlies. De tussenliggende tuniek is gepigmenteerd en vasculair en omvat van achteren het choroidea, ciliaire lichaam en iris.
De binnenste tuniek is nerveus en gevormd door het netvlies. De sclera stelt een expansie voor van de duralhuls van de oogzenuw, de choroïde is afgeleid van een uitzetting van pia-arachnoïde en de retina is in ontwikkeling een deel van de hersenen en is afgeleid van het diencephalon. Daarom is het netvlies een voorbeeld van een bewegend brein.
Tunieken van de oogbol:
Buitenste tuniek:
Het is vezelachtig en bestaat uit sclera en het hoornvlies (Fig. 9.21).
sclera:
De sclera is ondoorzichtig en vormt de achterste vijfzesde van de oogbol. Het is ononderbroken vooraan met het hoornvlies op de sclerosoomkruising en achter met de durale omhulling van de oogzenuw. Het is samengesteld uit een dicht vilt van collageenvezels.
Het voorste deel van sclera wordt door het bindvlies gezien als 'het wit van het oog'. Het uitwendige oppervlak van de sclera wordt bedekt door de fascia bulbi gescheiden door de episclerale ruimte en ontvangt de tendele inbrenging van zes extra-oculaire spieren.
De sclera wordt doorboord door de volgende structuren:
(a) De oogzenuw in het achterste deel samen met de centrale slagader en ader van het netvlies. De perforerende vezels van de zenuw maken het gebied meelevend; vandaar de lamina cribrosa sclerae die het zwakste deel van sclera is. Bij langdurige toename van de intraoculaire druk, zoals bij chronisch glaucoom, puilt lamina cribrosa naar achteren uit en vormt de cupping van de optische schijf;
(b) Achterste ciliaire vaten en zenuwen rond de oogzenuw;
(c) Ongeveer vier of vijf venae vorticosae doorboren de sclera in het midden tussen de bevestiging van de oogzenuw en de sclero-corneale overgang;
(d) Anterior ciliaire slagaders, afgeleid van de musculaire slagaders van vier recti, en waterige aderen die waterige humor afvoeren van de sinus venosus sclerae perforeren de sclera dichtbij de sclero-corneale overgang.
Functies van sclera:
(i) Het beschermt en onderhoudt de vorm van de oogbol;
(ii) Biedt de bevestiging van extra-oculaire spieren;
(iii) Ondersteunt de tussenliggende en innerlijke tunieken van de oogbal;
(iv) Behoudt de optimale intraoculaire druk van ongeveer 15 tot 20 mm Hg. Voor een goede veneuze terugkeer door de venae vorticosae moet de veneuze druk hoger zijn dan 20 mm Hg.
(v) Oogbol beweegt in de episclerale ruimte binnen de socket van fascia bulbi. Na operatieve verwijdering van het oog, fungeert de fascia bulbi als een socket voor prothese.

Hoornvlies:
Het is transparant, avasculair en vormt de voorste één-zesde van de oogbol. Het projecteert vooruit van de sclera, omdat het hoornvlies een segment van kleinere bol vertegenwoordigt. Uitwendig markeert een ronde groef, de sulcus sclerae, de kruising tussen het hoornvlies en de sclera. De dikte ervan is ongeveer 1 mm aan de omtrek en 0, 5 mm in het midden. Het is transversaal elliptisch op de anterieure surfage en rond op het achterste oppervlak.
Wanneer het hoornvlies meer gebogen is in de ene meridiaan dan in de andere, wordt de aandoening astigmatisme genoemd. Een lichte mate van astigmatisme is normaal aanwezig in de kindertijd en adolescentie, waarbij de kromming mogelijk groter is in de verticale dan in de horizontale meridiaan. De meeste breking door het oog vindt plaats, niet in de lens, maar aan de oppervlakte van het hoornvlies.
Voeding van het hoornvlies:
Omdat het hoornvlies avasculair is, krijgt het voeding door permeatie van drie bronnen:
(a) Lussen van capillairen aan de periferie van de conjunctivum-corneale overgang;
(b) Waterige humor vanuit de voorste kamer van het oog;
(c) De traansecretie verspreidt zich als een vloeistoffilm over het voorste oppervlak van het hoornvlies.
Structuur van het hoornvlies:
Het bestaat uit de volgende vijf lagen van buiten naar binnen (Fig. 9.22):
1. Hoornvliesepitheel:
Het bestaat uit niet-gekerstatiniseerd gelaagd squameus epitheel, meestal vijf cellen dik. Op de sclera-corneale overgang wordt het epitheel tien cellen dik en continu met de conjunctiva. De oppervlaktecellen presenteren microvilli die helpen bij het vasthouden van een ononderbroken film van traanvloeistof om het brekende oppervlak van het oog te vergroten. Het hoornvliesepitheel regenereert snel en wordt continu vervangen.
2. Bowman's membraan of anterieure begrenzende membraan:
Het vormt een acellulaire, dicht opeen gepakte laag van fijne collageenvezels en bedekt de onderliggende substantia propria.
3. Inhoudelijk: propria:
Het is samengesteld uit ongeveer 200 tot 250 op elkaar geplaatste afgeplatte lamellen. Elke lamel bevat bundels fijne collageenvezels die meestal evenwijdig aan elkaar en op het oppervlak van het hoornvlies lopen; de fibrillen lopen in opeenvolgende lamellen onder verschillende hoeken ten opzichte van elkaar.
Alle fibrillen hebben een uniforme grootte en zijn ingebed in een grondstof die rijk is aan chondroïtinesulfaat en keratosulfaat, waardoor het hoornvlies transparant wordt. De gemalen stof bevat ook fibroblastcellen met dendritische processen.
4. Descemet's membraan of posterieur beperkend membraan:
Het is een acellulaire, homogene, collageenachtige laag. Aan de periferie van het hoornvlies, verspreiden de collageenvezels zich achterwaarts om het trabeculaire weefsel te vormen dat de binnenwand van sinus venosus sclerae vormt en hechting verkrijgt aan het voorste oppervlak van de sclerale uitloper.
De ruimten tussen het trabeculaire weefsel bij de irido-corneale hoek communiceren de waterige humor van de voorste kamer van het oog naar de sinus venosus sclerae. Sommige vezels van het trabeculaire weefsel passeren mediaal naar de sclerale uitloper en zijn bevestigd aan de periferie van de iris als het pectinaat-ligament van iris.
5. Endotheel:
Het bestaat uit een enkele laag kubusvormige cellen die het achterste oppervlak van het hoornvlies bedekt, de ruimten van de iridocorneale hoek begrenst en wordt gereflecteerd op de voorkant van de iris.

Zenuwvoorziening van het hoornvlies:
Hoewel het hoornvlies avasculair is, bezit het een rijke sensorische zenuwtoevoer die afkomstig is van de oogheelkundige zenuw door de lange ciliaire zenuwen. De ciliaire zenuwen vormen vier opeenvolgende plexi als ze het hoornvlies bereiken:
(a) Een ringvormige plexus aan de rand van het hoornvlies;
(b) De zenuwvezels verliezen de myelineschede en vormen een propriale plexus in de substantia propria;
(c) Vezels van deze laatste vertakken onder het hoornvliesepitheel als sub-epitheliale plexus;
(d) Uiteindelijk penetreren de vrije zenuwuiteinden het epitheel en vormen ze intra-epitheliale plexus.
Eigenaardigheden van het hoornvlies:
1. De transparantie van het hoornvlies kan te wijten zijn aan de gladheid van het epitheel, de afwezigheid van bloedvaten, de uniforme organisatie van collageenvezels van de substantia propria en het type bodemsubstantie.
2. De allogene transplantatie van het hoornvlies zonder immunologische afstoting is opmerkelijk; het ontbreken van bloedvaten en afwezigheid van antigeen-presenterende cellen (APC's) zoals langerhans cellen van de huid voorkomen de afstoting van transplantaten.
Sclero-corneale overgang of limbus:
Binnen de substantie van sclera dichtbij de limbus en aan de periferie van de voorste kamer, bevindt zich een met endotheel bekleed rond kanaal bekend als de sinus venosus sclerae of het kanaal van Schlemm. In de sectie presenteert de sinus een ovale spleet, die in delen van zijn beloop dubbel kan zijn.
Grenzen van de sinus (Fig. 9.21)
De buitenste muur gemarkeerd door een sulcus in de sclera;
De binnenmuur:
(a) in het voorste gedeelte, gevormd door het trabeculaire weefsel dat is afgeleid van het membraan van de Descemet van het hoornvlies;
(b) In het achterste deel, gevormd door de sclerale uitloper, die een driehoekige projectie is van sclera die naar voren en naar binnen is gericht; het voorste oppervlak van de sclerale uitloper geeft hechting aan het trabeculaire weefsel en het achterste oppervlak verschaft oorsprong aan de ciliaris-spier.
Functie van de sinus:
1. Het verzamelt waterige humor uit de voorste kamer van het oog door de irido-corneahoek en de ruimten tussen het trabeculaire weefsel.
2. De waterlaag wordt afgevoerd van de sinus naar de voorste ciliaire aderen door middel van waterige aders die geen kleppen bevatten. Normaal bevat de sinus geen bloed; maar in veneuze congestie kan het worden gevuld met regurgitant bloed. Als de drainage wordt geblokkeerd, stijgt de intraoculaire druk, wat resulteert in een aandoening die bekend staat als glaucoom.
Gemiddelde tuniek:
Het is zeer vasculair en gepigmenteerd, en wordt vaak de uveal tractus genoemd, omdat na het ontleden van de sclera de blootgestelde tussenunit lijkt op de huid van een donkere druif die de geleiachtige inhoud van het oog omgeeft. De tussenliggende tuniek bestaat uit achter voorwaarts drie delen choroïde, ciliaire lichaam en iris.
choroid:
Het zit ingeklemd tussen de sclera en het netvlies, chocolade of donkerbruin van kleur. De choroidea lijnen de posterieure vijfzesde van de oogbol. Later wordt het doorboord door de oogzenuw, waar het hecht aan de sclera en continu is met de pia en arachnoidische maters.
Het buitenoppervlak is gescheiden van de sclera door de suprachoroïde lamina die is samengesteld uit een los netwerk van elastische en collageenvezels en wordt doorkruist door de lange achterste ciliaire vaten en zenuwen. Intern is het vaatvlies stevig gehecht aan de gepigmenteerde laag van het netvlies.
Structuur van choroid:
Van buiten naar binnen presenteert het de volgende lagen
1. Supra-choroïde lamina (lamina fusca) -vide supra.
2. Vasculaire lamina:
Het is een laag bloedvaten met verspreide pigmentcellen in het ondersteunende bindweefsel. De slagaders zijn afgeleid van korte posteriorale ciliaire slagaders en de aderen convergeren in kransen om vier of vijf vorticose-aders te vormen die de sclera doorboren en in de oftalmische aders uitmonden.
3. Capillaire lamina of choroïdopacillaire laag:
Het is een fijn netwerk van haarvaatjes die de buitenste drie of vier lagen retina voeden.
4. Basale lamina (membraan van Bruch):
Het is een dun, transparant membraan waaraan de gepigmenteerde laag van het netvlies stevig is gehecht.
Bij sommige dieren vormen gespecialiseerde cellen van het vaatvlies een reflecterend gebied dat bekendstaat als het tapetum dat 's nachts een lichtschaduw produceert in de ogen van sommige dieren.
Functies van de choroidea:
(a) Biedt voeding aan de buitenste lagen van het netvlies;
(b) Ondersteunt het netvlies, absorbeert het licht en voorkomt reflectie.
Ciliair lichaam (Fig. 9.23):
Het ciliaire lichaam strekt zich uit als een complete ring van het voorste deel van het choroïd in de ora serrata van het netvlies naar de periferie van de iris op de sclero-corneale overgang. Het biedt hechtingen aan het opschortende ligament van de lens en de perifere rand van de iris.

Omdat het dijbeen dikker is aan de voorzijde en dunner achter, is het ciliaire lichaam driehoekig in doorsnede, met de punt naar achteren gericht om het choroidea te verbinden. Het buitenoppervlak is in contact met de sclera. Het binnenoppervlak van het ciliaire lichaam is gericht naar het glasachtige lichaam achter en de zonulaire vezels van de lens (het opschortende ligament) aan de voorkant.
Het binnenoppervlak is deelbaar in twee ringzones: pars plicata in anterieur één derde en pars plana in posterior tweederde. De pars plicata presenteert zeventig tot tachtig ciliaire processen die meridionaal uitstralen van de periferie van de iris.
De verhogingen van de processen rusten in de groeven op het voorste oppervlak van het suspensieve ligament van de lens; de valleien tussen de processen geven hechtingen aan de zonulaire vezels van de lens die zich verder naar buiten uitstrekken om de pars plana binnen te dringen (Fig. 9.24).
De binneneinden van ciliaire processen steken uit in de periferie van de achterste kamer van het oog en scheiden waterige humor af. De pars plana of ciliaire ring is aan de rand begrensd door de ora serrata.
Een aantal lineaire richels die de bevestiging van lange zonulaire vezels verschaffen strekken zich radiaal naar buiten uit door de pars plana naar de toppen van ora serrata. Twee lagen epitheliale cellen van het netvlies worden verlengd over het binnenoppervlak van het gehele corpus ciliaris als pars ciliaris retinae en gaan dan verder op het achterste oppervlak van de iris; de diepere laag van ciliaire epitheel is gepigmenteerd.
Het korte voorste oppervlak of de basis van het driehoekige corpus ciliare geeft hechting aan de periferie van de iris nabij het midden ervan.

Structuur van het corpus ciliare:
Het bestaat uit stroma, ciliaris-spieren en een bilaminair epitheel dat het binnenoppervlak van het gehele ciliaire lichaam bedekt.
Ciliaire stroma omvat de supra-choroïde, vasculaire en basale laminae van de choroïde. Het bestaat uit losse fasciculi van collageenvezels die de ciliaire bloedvaten, zenuwen en ciliaris-spieren ondersteunen.
De slagaders zijn afgeleid van de lange posterior ciliaire slagaders die uiteenvallen in ingewikkelde gefenestreerde capillaire plexus in de ciliaire processen. Aan de periferie van de iris vormen de slagaders een belangrijke arteriële cirkel. De aderen komen samen met de vorticose-aders.
Ciliaris-spier (figuur 9.25):
Het is niet gestript en bestaat van buiten naar binnen uit drie sets vezels - meridionaal, radiaal en cirkelvormig. Alle vezels ontstaan vooraan vanaf het achterste oppervlak van de sclerale uitloper.
De meridionale vezels passeren posterieur door het stroma en zijn bevestigd aan de supra-choroïde lamina als terminale epichoroidale sterren.
De radiale of schuine vezels komen in de basis van ciliaire processen en verbinden zich onder een stompe hoek met elkaar.
De binnenste cirkelvormige vezels lopen omtrek na divergeren onder brede hoeken en vormen een soort sluitspier, dichtbij de omtrek van de lens.
acties:
Wanneer de ciliaris spier samentrekt, bewegen de supra-choroïde lamina en ciliaire processen naar voren. Uiteindelijk wordt het opschortende ligament van de lens ontspannen als gevolg van het vrijkomen van contactdruk. Dit maakt uitpuilen van de lens mogelijk om het oog aan te passen voor dichtbij zicht. Vandaar dat de ciliaris fungeert als spier van accommodatie.
De ciliaris-spier heeft geen tegenstander. Naarmate de spier ontspant, maakt de elastische terugslag van de supra-choroïde lamina de opschortende ligamentische spanning die resulteert in het afvlakken van de lens voor aanpassing van het gezichtsvermogen op afstand.
Zenuw voorraad:
Ciliaris-spier wordt geleverd door de parasympathische zenuwen. De pre-ganglionische vezels die voortkomen uit de Edinger-Westphal-kern van de oculomotorische zenuw in de middenhersenen worden doorgelaten naar het ciliaire ganglion. De post-ganglion-vezels bereiken de oogbol als korte ciliaire zenuwen.
Ciliair epitheel:
Het bestaat uit twee lagen epitheel, afgeleid van de twee lagen van de optische beker, voorbij de ora serrata van het netvlies. Cellen in de diepere laag zijn zwaar gepigmenteerd.
Iris:
De iris is een cirkelvormig, gepigmenteerd en samentrekkend diafragma dat is ondergedompeld in waterige humor tussen het hoornvlies en de lens. De perifere rand is bevestigd aan het voorste oppervlak van het corpus ciliare, en in de buurt van het midden ervan bevindt zich een cirkelvormige opening, de pupil.
De pupil vertegenwoordigt de rand van de ontwikkelende optische cup. De iris is geen afgeplatte schijf, maar lijkt eerder op een afgevlakte kegel die door de pupil is afgekapt, omdat het voorste oppervlak van de lens deze iets naar voren duwt.
Het voorste segment van de oogbol wordt door de iris verdeeld in voorste en achterste kamers die zijn gevuld met kamerwater en die via de pupil met elkaar communiceren (Fig. 9.21).
De voorste kamer wordt vooraan begrensd door het hoornvlies en achter door het voorste oppervlak van de iris en tegenover de pupil door het voorste oppervlak van de lens; de kamer is aan de periferie begrensd door de irido-hoek (hoek van filtratie) van waaruit waterig humeur wordt opgevangen in de sinus venosus door de ruimten van trabeculair weefsel (ruimtes van Fontana).
De achterste kamer wordt vooraan begrensd door het achterste oppervlak van de iris, en achter door de lens en zijn suspensieve ligament. Binnenuiteinden van ciliaire processen steken uit in de periferie van de achterste kamer en scheiden de waterige humor af.
Structuur van de iris (van vóór achteruit):
1. Het voorste oppervlak van de iris wordt niet bedekt door een duidelijk endotheel. Het presenteert opgravingen die bekend staan als crypten en een onregelmatige pony, de collarette die de bevestigingslijn van het pupilmembraan bij de foetus vertegenwoordigt. Een voorste grenslaag wordt gevormd door een laag vertakte fibroblasten en melanocyten en mengsels aan de periferie van de iris met het pectinaat-ligament dat afkomstig is van het Descemet-membraan van het hoornvlies.
2. Stroma van de iris (Fig. 9.25) - Het bevat collageenvezels, weefselruimtes ertussen, fibroblasten en melanocyten, bloedvaten en zenuwen, sfincter en dilatator pupillae spieren. De stromale ruimten staan in vrije communicatie met de vloeistof van de voorste kamer.

De sfincter pupillae is een ringvormige band van gladde spieren in het achterste deel van het stroma en omringt de pupil. De fusiforme spiercellen zijn gerangschikt in groepen en zijn ingesloten door een omhulsel van collageenvezels voor en achter.
Het is ontwikkeld uit het ectoderm en geleverd door de parasympathische vezels van de oogzenuwzenuw via de korte ciliaire zenuwen. De pupil is vernauwd wanneer de spier samentrekt. De iris trekt reflexmatig samen wanneer het licht het netvlies bereikt (pupilreflex) en tijdens aanpassing van het oog voor dichtbij zicht (accommodatie reflex).
Een druppel atropine in het oog verwijdt de pupil met verlies van accommodatie, omdat het medicijn de acties van sfincter pupillae en ciliaris spieren annuleert door de werking van acetyl choline op de effectorcellen te blokkeren.
De dilatator pupillae bestaat uit gladde spiervezels die stralen uit de collageen omhulsels die het achterste oppervlak van sfincter pupillen aan de rand van de pupil bedekken. De spier ligt onmiddellijk voor het gepigmenteerde epitheel en is afgeleid van de myo-epitheelcellen; vandaar dat de spier ectodermaal van oorsprong is.
De dilatator pupillae wordt geleverd door de sympathische zenuwen; de pre-ganglionische vezels zijn afgeleid van de laterale hoorncellen van T1- en T2-segmenten van het ruggenmerg, en de postganglionische vezels van de superieure cervicale sympathische ganglia bereiken de spier via de ciliaire zenuwen.
De iris ontleent zijn naam aan het Griekse woord regenboog vanwege zijn verschillende tinten van kleur. De kleur hangt af van de arrangementen en het type pigmenten en van de textuur van het stroma. Bij bruine iris zijn de pigmentcellen talrijk en bij blauwe iris is het pigment nauwelijks aanwezig. De blauwe kleur is te wijten aan diffractie en lijkt op de kleur van de blauwe lucht. Bij albino's is pigment afwezig in zowel het stroma als het epitheel en de roze kleur van de iris is te wijten aan bloed.
De pupil lijkt zwart, omdat de lichtstralen die door het netvlies worden gereflecteerd, worden gebroken door de lens en het hoornvlies en teruggaan naar de lichtbron. Een spleet in de iris die zich radiaal uitstrekt van de pupil staat bekend als het coloboma; het is een aangeboren afwijking en vertegenwoordigt een overblijfsel van choroïde spleet.
3. Gepigmenteerd epitheel:
Het is ook bekend als de pars iridis retinae die bestaat uit twee lagen gepigmenteerd epitheel en is afgeleid van het voorste deel van de ontwikkeling van optische cup. Het epithelium krult naar voren rond de rand van de pupil en vormt een zwarte, cirkelvormige pony.
Bloedtoevoer van de iris (fig. 9.26):
slagaders:
Aan de periferie van de iris (in plaats daarvan in het corpus ciliareus) wordt een belangrijke arteriële cirkel gevormd door de anastomose tussen de twee lange ciliaire slagaders achteraan en de anterieure ciliaire slagaders. Vanuit de hoofdcirkel passeer de bloedvaten centripetaal en anastomose dichtbij de pupilrand om een kleine arteriële cirkel te vormen die onvolledig kan zijn. De vaten zijn niet-gefenestreerd en zonder enige elastische lamina.

Aders begeleiden de slagaders en draineren in de vorticose aderen.
Zenuw voorraad:
Parasympathische zenuwen leveren de sluitspier pupillen, sympathische zenuwen leveren dilator pupillen en bloedvaten, en lange ciliaire zenuwen (oftalmisch) brengen sensorische vezels over.
Innerlijke tuniek:
Het wordt gevormd door een delicate neurale laag, het netvlies. In het achterste hoofdgedeelte van de oogbol bestaat het netvlies uit een gepigmenteerd buitengedeelte en een deel van het binnenste zenuwstelsel (neuro-retina), waarbij beide delen aan elkaar hechten.
Het achterste deel van het netvlies, ook wel het optische deel van het netvlies genoemd, strekt zich uit van de bevestiging van de oogzenuw (optische schijf) achter de gekantelde rand, oftewel een serrata, aan de voorkant waar het neuro-netvlies stopt.
De ora serrata ligt aan de periferie van het corpus ciliare. Voorbij de ora serrata strekt het bilaminaire membraan van het niet-nerveuze deel van het netvlies zich naar voren uit over het ciliaire lichaam en de iris die respectievelijk pars ciliaris en pars iridis-netvlies vormen.
Extern is het optische deel van het netvlies nauw verbonden met de basale lamina van het vaatvlies; intern wordt het gescheiden van het glaslichaam door het hyaloïde membraan. Het gepigmenteerde deel van het netvlies is ontwikkeld uit de buitenste wand van de optische kom en het nerveuze deel ervan uit de binnenwand van de optische beker.
In het late foetale leven wordt de intra-retinale ruimte weggevaagd en worden de twee delen van het netvlies gefuseerd. Bij het losmaken van de retina wordt de gepigmenteerde laag gescheiden van de neuro-retina en dit is de algemene oorzaak van gedeeltelijke blindheid.
De neuro-retina bevat in lagen een groot aantal onderling verbonden sensorische neuronen,
interneuronen, ondersteunende neurogliacellen en bloedvaten. De buitenste laag bevat fotoreceptorcellen, staafjes en kegeltjes. De staven zijn laagdrempelig en gevoelig voor zwak licht (scotopisch zicht).
De kegels hebben een hogere drempelwaarde (fotopisch zicht) en hebben betrekking op helder licht en kleurenvisie. Fotoreceptorcellen van het netvlies ontvangen een omgekeerd beeld van het object. Er is nog steeds onenigheid over hoe men zich aanpast aan de inversie van het netvliesbeeld. Licht moet door alle netvlieslagen heen voordat het Rods en Cones bereikt.
Van de fotoreceptoren hebben de eerste neuronen hun cellichamen in de bipolaire cellen van het netvlies. Ze maken synapsen met de tweede neuronen in de ganglioncellen van het netvlies, waarvan de axonen naar het laterale geniculaire lichaam passeren, vandaar, na relais, de derde neuronen naar de primaire visuele cortex van de occipitale lob door middel van optische straling.
Twee gebieden van het netvlies verdienen speciale vermelding, optische schijf en macula lutea.
Optische schijf:
Het is een cirkelachtig bleek gebied waaruit de oogzenuw begint en meet ongeveer 1, 5 mm in diameter. De schijf bevindt zich enigszins mediaal en superieur aan de achterste pool van de oogbol. Het ligt boven de lamina cribrosa van de sclera.
De optische schijf is verstoken van staafjes en kegeltjes; daarom is het ongevoelig voor licht dat de dode hoek vormt. Een normale schijf vertoont een variabele depressie in het midden die bekend staat als de fysiologische cup. De centrale vaten van de retina passeren de schijf dichtbij het midden ervan.
Het oedeem van de schijf dat bekend staat als het papiloedeem (gesmoorde schijf) kan worden waargenomen door de oftalmoscoop bij verhoogde intra-craniale druk als gevolg van compressie op de centrale ader van het netvlies, terwijl de laatste door de subarachnoïdale ruimte rond de oogzenuw gaat.
Macula lutea:
Het is een gelig gebied aan de achterste pool van het oog, ongeveer 3 mm lateraal aan de optische schijf. De gele kleur is te wijten aan de aanwezigheid van xanthofylpigment. Macula meet ongeveer 2 mm horizontaal en 1 mm verticaal.
De macula vertoont een centrale verlaging, de fovea centralis, waarvan de onderkant bekend staat als de foveola, die avasculair is en wordt gevoed door de choroïde. De fovea centralis, ongeveer 0, 4 mm in diameter, is het dunste deel van het netvlies, omdat de meeste lagen van het netvlies, met uitzondering van de kegeltjes, naar de buitenrand worden verplaatst.
Alleen kegels zijn aanwezig in de fovea; in elk menselijk netvlies zijn foveale kegels ongeveer 4000. Hier is elke kegel verbonden met slechts één ganglioncel door één dwergbipolaire cel. Daarom houdt de fovea zich bezig met een discriminerend zicht.
Structuur van het optische deel van het netvlies:
Conventioneel wordt beschreven dat het netvlies de volgende tien lagen van buiten naar binnen bezit (Fig. 9.27).
1. Gepigmenteerd epitheel;
2. Laag van de buitenste en binnenste segmenten van de stangen en kegels;
3. Extern beperkend membraan:
Het wordt gevormd door de nauwe overgangen tussen de buitenste geëxpandeerde uiteinden van retinogliacellen (de cellen van Muller) waarop binnenste segmenten van staven en kegeltjes rusten;
4. Buitenste nucleaire laag:
Het bevat de cellichamen van Rods and Cones en hun binnenste vezels;
5. Outerplexiforme laag:
Hier verstevigen staafbollen en kegelspieren synaps met bipolaire en horizontale cellen;
6. Binnenste nucleaire laag:
Het bevat de cellichamen van horizontale cellen in de buitenste zone, Amacrine cellen in de binnenste zone; en de Bipolaire en Müller-cellen in de tussenliggende zone;
7. Binnenste plexiform laag:
Het wordt ingenomen door de synapsen tussen de Bipolaire, Amacrine en Ganglion-cellen;
8. Ganglion cellenlaag:
Het bevat de cellichamen van ganglioncellen;
9. Zenuwvezellaag (stratum opticum):
Het wordt gevormd door de niet-gemyeliniseerde axonen van ganglioncellen; de vezels convergeren naar de optische schijf, doorboren de retina, choroïde en lamina cribrosa van sclera waar ze hun myeline schede ontvangen en de oogzenuw vormen;
10. Intern beperkend membraan, wordt gevormd door junctionele complexen van geëxpandeerde binnenste uiteinden van Muller's cellen op een vitaal oppervlak.
Functioneel maken drie sets van neuronen synaptische contacten in de retina in longitudinale kolommen. Deze zijn van buiten naar binnen genoemd als volgt: -
(a) Fotoreceptorstaven en kegelcellen en hun processen strekken zich uit van de tweede tot de vijfde laag van het netvlies;
(b) Bipolaire cellen met hun dendrieten en axons bezetten de 5e tot 7e laag;
(c) Ganglioncellen en hun axonen worden in de 8e en 9e lagen geplaatst. De longitudinale neuronale kolommen worden horizontaal geïntegreerd door de horizontale cellen en Amacrine cellen. Al deze neuronen worden ondersteund door de retinoglia-cellen (Mullers cellen), waarvan de buitenste en binnenste geëxpandeerde uiteinden zijn verenigd door nauwe overgangen om respectievelijk de externe en interne beperkende membranen te vormen.

Pigmentepitheel:
Het bestaat uit een enkele laag kubusvormige cellen die op de basale lamina van de choroïde rusten. Het cytoplasma bevat, naast organellen, melaninepigmenten in melanosomen. De cytoplasmatische infoldings beïnvloeden de basale zone van de cellen; hun apicale zone is voorzien van microvilli die uitsteken tussen de buitensegmenten van stangen en kegels.
functies:
(a) Microvilli van de pigmentcellen fagocyteren versleten lamellen van de buitenste segmenten van staven en kegeltjes en degraderen ze door lysosomale werking. Aldus helpen de pigmentcellen bij de omzet van staaf- en kegel-fotoreceptorcomponenten;
(b) Absorbeer de lichtstralen en voorkom achterwaartse reflectie;
(c) Zorg voor voeding aan de avasculaire zone van de buitenste drie of vier lagen retina door diffusie vanuit de aangrenzende capillaire plexus van de choroïde;
(d) De pigmentcellen werken door hun nauwe verbindingen met elkaar als bloed-retinale barrière om een speciale ionische omgeving van het netvlies te handhaven, samen met transport van groeifactoren. De barrière voorkomt de invoer van immunologisch competente lymfocyten in de retina.
Hengels en kegels:
Staven en kegeltjes zijn langwerpige fotoreceptoren die gepolariseerd en gesegmenteerd zijn in subregio's met verschillende functionele rollen. Elke fotoreceptor bestaat uit een buitenste segment, een verbindingssteel en een binnenste segment, een cellichaam met een vezel en een synaptische basis.
Het buitenste segment van een staaf is cilindrisch en dat van een kegel is kort conisch. Elk buitensegment bevat talrijke afgeplatte vliezingsschijven die haaks op de lange as van de cel zijn georiënteerd. Alle schijven van Cone behouden hun continuïteit met het celmembraan. De meeste schijven van een staaf hebben geen hechting aan het celmembraan. De visuele pigmentmoleculen zijn opgenomen in de schijven.
De smalle verbindingssteel tussen buitenste en binnenste segmenten is een cytoplasmatische brug die een cilium omsluit.
Het binnenste segment is verdeeld in een buitenste ellipsoïde zone en een binnenste myoid zone. De ellipsoïde is gevuld met mitochondriën en de myoïde bevat Golgi-apparatuur en endoplasmatisch reticulum. De cytoplasmatische organellen van de myoïde synthetiseren nieuwe fotoreceptor-eiwitten die naar de vliezige schijven worden getransporteerd.
De schijven worden verplaatst naar het choroidea wanneer de nieuw gevormde schijven worden toegevoegd. Uiteindelijk worden de schijven van de uiteinden van de buitensegmenten afgeworpen en worden ze in de pigmentcellen opgenomen voor verwijdering.
De visuele pigmenten van Rods en Cones zijn samengesteld uit een specifiek eiwit, een opsine, dat is gebonden aan een chromatofoor, het retinaldehyde, met een speciale configuratie. Photopigment of the Rods staat bekend als rodopsin (visueel paars), die de sensoren zijn van zwart, grijs en wit. De kegeltjes bevatten drie fotopigmenten - blauw, groen en rood, waarbij elke kegel één pigment bevat.
Alle vier fotopigmenten bezitten de 11-cm retinaldehyde als de chromatofoor en zijn verenigd tot vier verschillende opsins. De actie van licht is om het retinaldehyde van de 11-c / s naar de all-trans-configuratie te isomeriseren. De chemische stappen die worden getriggerd door de lichtgolven in de fotopigmenten zijn verantwoordelijk voor het ontstaan van het receptorpotentieel in de staafjes en kegeltjes.
De axonachtige vezel is een cytoplasmatische verlenging van Rod of Cone, die een cellichaam met zijn kern omvat. Elke vezel eindigt in een gespecialiseerd synaptisch lichaam, dat in synaptisch contact komt met de zenuwvezels van bipolaire en horizontale cellen.
Het synaptische lichaam van een kegel is vlak en staat bekend als een steeltje, dat van een staaf wordt een bol genoemd omdat het klein en afgerond is. De rod-spherule die synaptische linten bevat, vertoont een oppervlaktedepressie die contact maakt met de dendrieten van bipolaire cellen en processen van horizontale cellen. De kegel pedicles presenteert drie soorten synaptische contacten:
(a) Het draagt een aantal depressies, die elk een synaptische triade vormen met drie neurietterminals; twee diepgeplaatste processen zijn afgeleid van horizontale cellen en een dendritische terminal van midget bipolaire cellen;
(b) Platte oppervlakken tussen depressies vormen synapsen met platte bipolaire cellen;
(c) De periferie van de puntstukjes vormt contacten met de staafbollen. De 'ON'-bipolaire cellen maken synaptische triaden in de depressieve gebieden van staafbolletjes en kegelstammen, terwijl' UIT'-bipolaire cellen in contact komen met de platte oppervlakken van de kegelsteeltjes.
In de rustende (donkere) staat is er spontane afgifte van neurotransmitter van de staven en kegeltjes naar de bipolaire cellen, vanwege een gestage instroom van ionen door de natriumkanalen in het membraan vanwege de beschikbaarheid van voldoende cGMP.
Maar in aanwezigheid van licht wordt cGMP uitgeput door gecompliceerde moleculaire gebeurtenissen van transductie zodat de natriumpoort gesloten is. Dit veroorzaakt hyperpolarisatie van staven en kegeltjes die de spontane afgifte van neurotransmitter over het synaptische gebied van de buitenste plexiforme laag remmen.
Er zijn ongeveer 120 miljoen staven, 7 miljoen kegeltjes in het netvlies van elk oog en ongeveer 1 miljoen Ganglion-zenuwvezels om elke oogzenuw te vormen. Therefore, convergence of retinal information takes place from the receptors to the Ganglion cells.
Rods are numerous in the peripheral part and absent at the fovea centralis; Cones are concentrated in the central part and at the fovea centralis Cones only are present, having about 4000 in number. Both Rods and Cones are absent at the optic disc (Blind spot). In the peripheral retina, about 200 Rods converge on one Bipolar cell and as many as 600 Rods converge through intemeurons on one Ganglion cell.
Bipolar cells:
The Bipolar cells are divided into two principal groups, Cone and Rod bipolars.
Cone bipolars:
These consist of three major types: midget, blue and diffuse.
The midget bipolar possesses a small body, as the name implies. The single dendrite of each midget cell makes synapses with the pedicle of only one cone, which may be invaginating triadic type or flat type; the former represents the 'ON' bipolar and the latter 'OFF' bipolar cell.
The axon arising from the other pole of the cell enters the inner plexiform layer and synapses with the dendrites of a single ganglion cells and with the neurites of different classes of amacrine cells.
The 'ON' cell synapses in the middle stratum and 'OFF' cell in the outer stranum of inner plexiform layer. The 'ON' cells respond by depolarisation and 'OFF' cells by hyperpolarisation The midget bipolars are connected with either red- or green- sensitive cones.
Blue cone bipolars are slightly larger than the midget cells and establish similar connection between a single cone and a single ganglion cell. Such unitary one—to—one channel involving midget and blue cone bipolar cells conveys trichromatic information with high visual acuity.
Diffuse cone bipolars are fairly large and possess wide receptive areas connecting with 10 or more cones. They are concerned with luminosity rather than the colour. There are six distinct diffuse cone bipolars: three are 'ON' type and three 'OFF' type.
Rod bipolar:
These are connected by branched dendrites with numerous rod spherules forming invaginating triad synapses and as such all belong to 'ON' cells. The axon of each rod bipolar reaches the inner stratum of inner plexiform layer and synapses indirectly with the ganglion cells through the amacrine cells.
Horizontal cells:
These are situated in the outer zone of inner nuclear layer of retina. The horizontal cells are inhibitory interneurons using GAB A as a neuro-transmitter. Their dendrites and axons extend within the outer plexiform layer, and make synapses with a number of rod spherules and cone pedicles depending on the position of retina and also come in contact with the adjacent horizontal cells via gap junction.
The participation of two axon terminals of horizontal cell centering the single dendrite of rod or cone bipolar cells in the synaptic triad helps neural sharpening of 'ON' bipolar cells by lateral inhibition.
Amacrine cells:
These cells do not possess typical axon, hence the name. But their dendrites function both as axons and dendrites and are involved in both incoming and outgoing synapses. The cell bodies of amacrine cells usually occupy the inner zone of the inner nuclear layer, but some are displaced to the outer aspect of ganglion cell layer.
Their neurites spread in the three strata of inner plexiform layer and are connected with the axons of bipolar cells, dendrites of ganglion cells and the processes of other amacrine cells. One class of amacrine cells (A-II) transmits signals from rod bipolars to the ganglion cells.
According to their neurotransmitter contents, the amacrine cells are classified as glycinergic, GABA-ergic, cholinergic etc. But their functions are not yet explored. However, the following functions deserve special consideration:
ik. They modulate the photoreceptive signals;
ii. Act as essential element in transmitting signals from rod bipolars to ganglion cells;
iii. Maintain a balance of illumination sensibility between the two halves of retina;
iv. Amacrine cells are possibly connected with the retino-petal fibres which enter the retina through the optic nerve. It is not unlikely that they may arise from reticular neurons of the brain stem and are concerned with the arousal or inhibitory response of vision, since the retina is developmentally a moving brain.
Inner plexiform layer:
As described earlier, the inner plexiform layer is divisible into three strata:
(a) Outer 'OFF' stratum consists of 'OFF' bipolar cells connecting with the dendrites of ganglion cells and neurites of amacrine cells;
(b) Middle 'ON' stratum where 'ON' bipolar cells synapse with the dendrites of ganglion cells and neurites of amacrine cells;
(c) Inner Rod stratum where Rod bipolars make synapses with the neurites of displaced amacrine cells.
Mechanism of 'ON' and 'OFF' bipolar response:
ik. In het donker wordt de neurotransmitter maximaal vrijgegeven van de synaptische knooppunten van staven en kegels. Dus, in het donker depolariseert de neurotransmitter de 'OFF' bipolaire cellen en hyperpolariseert de 'ON' bipolaire cellen.
ii. Tijdens de belichting van het netvlies neemt het neurotransmitterniveau af en dat resulteert in hyperpolarisatie van de 'OFF'-cellen en depolarisatie van de' ON'-cellen. Uiteindelijk geven de 'ON'-cellen de neurotransmitter vrij aan hun axon-aansluitpunten, terwijl de hypergepolariseerde' OFF'-cellen de afgifte stoppen.
Retino-gliacellen van Muller:
Hun cellichamen liggen in de binnenste nucleaire laag, en hun buitenste en binnenste cytoplasmatische processen vormen de externe en interne beperkende membranen.
De Muller-cellen ondersteunen niet alleen de neuronen van het netvlies, ze slaan glycogeen op in hun cytoplasma dat door omzetting in glucose een kant en klare energiebron is voor complexe biochemische activiteiten in het netvlies.
Ganglion cellen:
Het aantal Ganglion-cellen in elk menselijk netvlies is ongeveer 1 miljoen. Deze zijn in principe van twee typen, dwerg (β) cellen en parasol (α) cellen.
Midget ganglioncellen op het maculaire gebied verbinden zich met een enkele dipolaire of blauwe kegel bipolaire dij en een enkele kegelsteel, en hebben betrekking op scherpte van zicht en kleurdiscriminatie.
Parasol ganglioncellen gelokaliseerd aan de periferie van het netvlies hebben uitgebreide ontvangende velden die de invoer ontvangen van de diffuse kegel bipolairen en staaf bipolairen via de amacrine cellen en primair signaalveranderingen van verlichting.
De axonen van dwergganglioncellen projecteren in het parvocellulaire gedeelte en die van parasolcellen in het magnocellulaire deel van het laterale geniculaire lichaam; vandaar respectievelijk P- en M-cellen.
Sommige van de subklassen van beide typen ganglioncellen reageren op het begin van de verlichting en worden 'AAN'-cellen genoemd, en de andere worden geactiveerd door compensatie van uitlichting en worden' UIT'-cellen genoemd. Sommigen worden echter tijdelijk geactiveerd door zowel het begin als het uitschakelen van de verlichting en worden aangeduid als 'AAN-UIT'-cellen.
De dendrieten van 'ON'-ganglioncellen maken synapsen met de middelste stratum, ' OFF'-cellen met de buitenste laag en die van 'ON-OFF'-cellen met zowel' ON'- als 'OFF'-lagen van de binnenste plexiformlaag.
Bloedvoorziening van het netvlies (fig. 9.28):
De binnenste zes of zeven lagen van het netvlies worden geleverd door de centrale ader; de buitenste drie of vier lagen zijn avasculair en krijgen voeding door diffusie vanuit de capillaire lamina van de choroïde. De centrale slagader, een aftakking van oogheelkundig oog, passeert de lamina cribrosa in de oogzenuw en bij het bereiken van de optische schijf verdeelt deze zich in een bovenste en een onderste tak.
Elk geeft nasale en tijdelijke vertakkingen af. Deze vier takken zijn eindaders en leveren hun eigen kwadranten van het netvlies. Binnen de kwadranten splitsen vertakkingen van de retinaslagader zich dichotom en de twee rami divergeren onder een hoek van 45 ° tot 60 °. Elke blokkering van een retinale slagader wordt gevolgd door verlies van gezichtsvermogen in het overeenkomstige deel van het gezichtsveld.

De kronkels van de retinale venen convergeren op de optische schijf om de centrale ader van het netvlies te vormen en uiteindelijk in de caverneuze sinus af te tappen. Het oftalmoscopisch onderzoek onthult dat de retinale slagaders voor de retinale aderen kruisen.
De brekende media van het oog:
Het brekingsapparaat van het oog omvat het hoornvlies, de waterige humor, de lens en het glaslichaam. Ongeveer tweederde van de breking van licht vindt plaats aan de voorkant van het hoornvlies op de kruising van lucht en hoornvliesepitheel.
Waterige humor:
Het vult de voorste en achterste kamers van het oog en is samengesteld uit ongeveer eiwitvrij plasma. Het bevat glucose, aminozuren, een beetje hyaluronzuur, een hoge concentratie vitamine С en zorgt voor de ademhalingsgassen.
De humor in water wordt gevormd door een actief proces en door diffusie van de capillairen van de ciliaire processen en wordt aanvankelijk verzameld in de achterste kamer. Vandaar dat het via de pupil in de voorkamer verschijnt. Omdat de pupil nauw wordt toegepast op het voorste oppervlak van de lens, kan het fluïdum van de achterste naar de voorste kamer stromen, maar niet in de omgekeerde richting.
Bij het bereiken van de irido-hoornvlieshoek vinden de vloeistoffen zijn weg in de sinus venosus sclerae (kanaal van Schlemm) door de met endotheel omgeven ruimten van het trabeculaire weefsel (Spaces of Fontana). Uiteindelijk wordt het via de waterader naar de voorste ciliaire aderen afgevoerd. Enige hoeveelheid van de vloeistof wordt ook geabsorbeerd door het voorste oppervlak van de iris in de iridiale veneuze plexus.
functies:
(a) Het levert voeding aan het hoornvlies en de lens.
(b) het handhaaft de intraoculaire druk; de normale druk is ongeveer 15-20 mm Hg. en wordt berekend door tonometrie uit metingen van de indrukbaarheid van het verdoofde hoornvlies.


Lens:
De lens is een transparant, biconvex, flexibel lichaam dat ingrijpt tussen de iris vooraan en het glasachtige lichaam erachter. Het presenteert anterior en posterior oppervlakken, gescheiden door een afgeronde rand, de evenaar. Het achterste oppervlak is meer convex dan het voorste oppervlak, en het rust op een hyaloïde fossa van het glaslichaam.
De middelpunten van beide oppervlakken zijn bekend als de polen en de lijn die de voorste en achterste polen verbindt vormt de as van de lens. -Het is verankerd aan het ciliaire lichaam door de zonulaire vezels, die zich vermengen met de lenscapsule rond de evenaar en het opschortende ligament van de lens vormen.
De gemiddelde diameter van de lens is ongeveer 1 cm. Het draagt ongeveer 15 dioptrie bij tot het totaal van 58 dioptrische kracht van het oog. Het voordeel van de lens ten opzichte van andere brekende media is dat deze de dioptrische kracht kan wijzigen voor dichtbij of veraf zien door de kromming van het vooroppervlak aan te passen.
De lens absorbeert veel ultraviolet licht. Naarmate de leeftijd vordert, wordt de lens steeds geeler en harder. Als gevolg hiervan wordt de kracht van accommodatie voor dichtbij zien verminderd waardoor presbyopie wordt geproduceerd. Dit defect kan worden gecorrigeerd door het gebruik van convexe brillen. De dekking van de lens staat bekend als cataract.
Structuur van de lens (Fig. 9.29):
Het bestaat uit een capsule, anterieure epitheel en lensvezels.
Lens capsule:
Het is een transparant, elastisch basismembraan met een overvloed aan reticulaire vezels en omhult de gehele lens. Het wordt gevormd door de epitheellenscellen, bevat gesulfateerd glycosaminoglycan en kleurt schitterend volgens de PAS-techniek. Op de evenaar van de lens is de capsule gefuseerd met de vezels van de ciliaire zonule.

Anterior epithelium:
Onder de capsule is het voorste oppervlak van de lens bekleed met een enkele laag laag kubisch epitheel. Richting de evenaar van de lens, verlengen de epitheelcellen zich en differentiëren ze tot lensvezels die meridionaal draaien en het grootste deel van de lenssubstantie vormen.
Lensvezels [Fig. 9.30 (a), (b), 9.31]:
De gelamineerde structuur van de lens is het gevolg van de continue toevoeging van vezels in het gebied van de evenaar en dit proces gaat door gedurende het hele leven. Tijdens de transformatie van lenscellen in de lensvezels verliezen de oude vezels in het midden hun kernen en de nieuwe vezels aan de omtrek bezitten afgeplatte kernen.
Vandaar dat het hardere centrale deel van de lens bekend staat als de kern en het periferaal zachtere deel de cortex vormt. De kernachtige en niet-genucleëerde lensvezels blijven in leven en bevatten longitudinaal geplaatste microtubuli en karakteristieke kristallijne eiwitten.
In afwezigheid van kernen wordt de eiwitsynthese van de vezels gehandhaafd door langlevend mRNA. Het aantal lensvezels bij volwassenen is ongeveer 2000. Op de doorsnede is elke vezel een hexagonaal prisma.

Voorzieningen van lensvezels:
De lens is ontwikkeld uit een lensblaasje tijdens de zesde week van het embryonale leven door de invaginatie van het oppervlakte-ectoderm. Daarna gaat het vesicle terug van het oppervlak en ligt in de holte van de optische beker.
De voorste wand van de blaar bestaat uit een enkele laag kubusvormig epitheel. De cellen van de achterwand van de vesikel strekken zich van achteren naar voren uit en worden omgezet in de primaire lensvezels; deze vezels vernietigen uiteindelijk de holte en ontmoeten de voorste wand.
De vernietiging is voltooid tegen de zevende week. De cellen van de voorste wand van de vesikel blijven echter intact, prolifereren en migreren naar de evenaar van de lens, waar ze langwerpig zijn en differentiëren tot de secundaire lensvezels.

De primaire lensvezels die postero-anterieur groeien, zijn gerangschikt in vellen die op beide oppervlakken van de lens worden uitgedrukt als Y-vormige hechtingen. De voorste Y staat rechtop, terwijl de achterste Y is omgekeerd. De secundaire lensvezels strekken zich gebogen uit vanaf het hechtmateriaal op het vooroppervlak naar dat op het achterste oppervlak.
De opstellingen van de vezels zijn zodanig dat die welke ontstaan uit het midden van Y op één oppervlak eindigen in de uiteinden van Y op het tegenoverliggende oppervlak, en omgekeerd. De Y-vormige hechtingen zijn in vivo te zien met een spleetlampmicroscoop.
Glasachtig lichaam:
Het is een transparante, gelatineuze massa die de achterste viervijfde van de oogbal vult. Het is samengesteld uit 99% water met enkele zouten en bevat een netwerk van collagene fibrillen en een mucopolysaccharide, het hyaluronzuur.
Een smal hyaloidekanaal strekt zich voorwaarts door het lichaam uit van de optische schijf naar het midden van het achterste oppervlak van de lens. Het kanaal is bezet in het foetale leven door de hyaloidader (een voortzetting van de centrale slagader van het netvlies), die normaal ongeveer zes weken voor de geboorte verdwijnt.
Het glaslichaam is omhuld door een gevoelig en transparant hyaloid membraan, dat is bevestigd aan het ciliaire epitheel en ciliaire processen en aan de rand van de optische schijf. Anteriorly, vormt het membraan een depressie, de hyaloïde fossa, waarop het achterste oppervlak van de lens rust.
Voor ora serrata wordt het hyaloïde membraan verdikt door de introductie van radiale vezels om de ciliaire zonule te vormen. In dit gebied vertoont het membraan een reeks groeven waarin de ciliaire processen zijn ondergebracht.
De ciliaire zonule splitst zich in twee lagen - de achterste laag bedekt de vloer van de hyaloïde fossa; de voorste laag scheidt zich af in zonulaire vezels die aan de lenscapsule voor en achter de evenaar zijn bevestigd.
De zonulaire vezels vormen gezamenlijk het opschortende ligament van de lens en zijn aan de omtrek bevestigd aan de groeven tussen de ciliaire processen en strekken zich verder naar buiten uit als lineaire richels naar de toppen van de ora serrata.